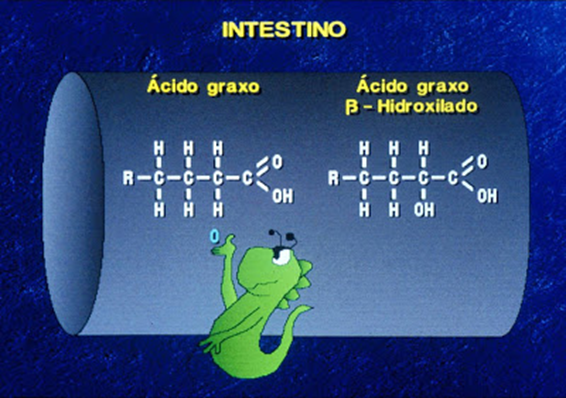
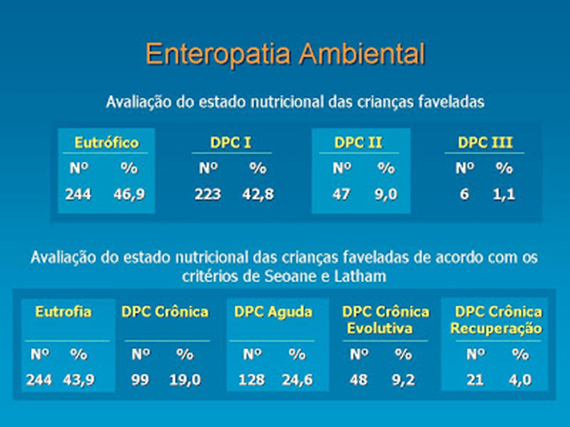
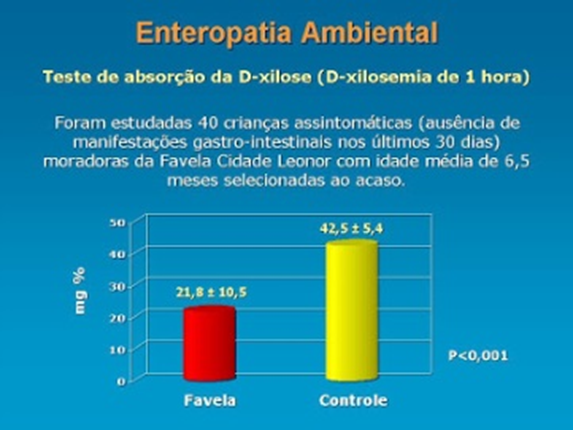
A
história da realização de um Pós-Doutorado no North Shore University Hospital,
Cornell University, Nova Iorque: pessoal e científica
A
realização do projeto de Pesquisa Experimental: “Absorção de
Macromoléculas Intactas Pelo Jejuno De Ratos in vivo: Influência Dos Sais Biliares Taurocolato,
Colato e Deoxicolato”
Material e Métodos
1. Perfusão
Intestinal
Ratos
machos, cêpa Wistar, pesando entre 100 e 150 gramas foram utilizados. Os ratos
foram alimentados com dieta padrão até a noite anterior ao experimento.
Os
ratos foram anestesiados pela via intra-peritoneal com uretano à dose de 1,2
g/kg de peso. A cavidade abdominal foi aberta por uma incisão médio-ventral e o
duodeno identificado. Uma pequena incisão foi realizada na porção média do
duodeno e um tubo de polietileno foi inserido através de uma incisão até
atingir o ligamento de Treitz. A cânula foi então fixada no lugar por meio de
uma ligadura. Um segmento de jejuno, medindo aproximadamente 30-40 cm, foi
utilizado para perfusão e a extremidade distal deste segmento também foi
canulada, a cânula foi fixada por uma ligadura. O segmento distal assim
preparado para a perfusão intestinal foi delicadamente lavado com solução
fisiológica isotônica evitando-se distensão da alça intestinal a ser perfundida
(Figura 11).
Figura 11-
Início do procedimento cirúrgico para implantação das cânulas; às vezes ocorria
algum acidente de trabalho como mostra a foto.
A
cânula proximal foi acoplada a uma bomba de perfusão peristáltica marca Harvard
modelo 1220 e o segmento jejunal devidamente preparado foi perfundido durante
60 minutos a um ritmo de 0,21-0,24 ml/min com uma solução isotônica (280
mOsm/l) de glicose e cloreto de sódio (pH 6,9) contendo 600 mg% de polietileno-glicol
(PEG) e 0,5 g% de Horseradish peroxidase (HRP) (Tipo II Sigma Chemical
Co., St Louis, M) como marcador proteico macromolecular. A esta
solução de perfusão foi acrescentada, de forma independente para cada sessão do
experimento, o sal sódico de um dos seguintes ácidos biliares: Taurocólico
(TCh) (Sal biliar primário e conjugado), Cólico (Ch) (Sal biliar primário e
desconjugado) e Deoxicólico (DCh) (Sal biliar secundário e desconjugado) a uma
concentração de 0,5 mM. Ratos Controles (C) foram perfundidos com uma
solução isenta de sais biliares. Após a adição dos sais biliares o pH da
solução foi reajustado para o valor de 6,9 e a osmolaridade permaneceu
inalterada (Figura 12).

Figura
12- Os ratos em plena seção de perfusão intestinal após terem sido
apropriadamente preparados para tal com as cânulas proximais acopladas às
bombas de perfusão. Cada experimento envolvia 12 ratos em um mesmo
procedimento.
A
concentração 0,5 mM dos sais biliares foi escolhida por ser aquela que
fisiologicamente se encontra no lúmen intestinal dos ratos adultos. Foram
estudados os fluxos de água por meio da determinação das alterações
intra-luminais de um marcador não absorvível, o PEG, durante
o experimento, expressos segundo a seguinte relação PEG inicial/PEG
final, ou seja determinando-se o quociente entre a concentração do PEG
na solução infundida e a concentração do PEG na solução
coletada da perfusão na cânula distal. O transporte do sódio foi analisado
determinando-se a concentração do sódio perfundido em aparelho de fotometria de
chama (IL Modelo 143) e a absorção de glicose pelo método da glicose oxidase.
Os cálculos finais dos transportes de sódio e glicose foram determinados
aplicando-se a seguinte fórmula: T= C inicial – (C final x PEGr) x
Ritmo de perfusão (ml/min) x 1000/ Comprimento de alça intestinal perfundida
(cm), aonde: C inicial representa a concentração da
substância na solução de perfusão e C final a concentração da
substância no perfundido e PEGr o valor numérico da
relação PEG inicial/PEG final. Valores finais positivos
indicam absorção de uma determinada substância e negativos indicam secreção.
2. Análise
Bioquímica da Absorção de HRP
A
absorção de HRP desde a luz intestinal para a circulação sistêmica foi
monitorada enzimaticamente. Ao cabo dos 60 minutos do período experimental,
sangue da aorta abdominal foi cuidadosamente obtido, evitando-se hemólise. Este
procedimento foi feito empregando-se uma agulha número 22 e seringa
heparinizada da qual se retirou o êmbulo para que o sangue pudesse fluir
espontaneamente. A agulha era então removida e permitia-se que o sangue fluísse
da seringa para um tubo de ensaio conservado em um balde com gelo. O plasma era
separado por centrifugação a 500g durante 10 minutos e posteriormente
clarificado por uma segunda centrifugação sob idênticas condições (Figura 13).
Figura 13- Mary Ann fazendo a coleta
de sangue da aorta do rato ao término do experimento de perfusão para fazer a
dosagem sérica da HRP. Estou ao seu lado e há um buraco esbranquiçado na minha
calça, produto de manipular substâncias corrosivas no laboratório.
A
atividade sérica de HRP foi determinada utilizando-se orto-dianisidina (corante)
como doadora de electron (Worthington Biochemical Corp., Freehold, NJ). Os
resultados foram expressos como micromoles de peroxidase decomposta/min/ml/cm
de jejuno perfundido e corrigido para um padrão de ritmo de perfusão de 0,2
ml/min. A reação enzimática obedece ao seguinte princípio básico: a peroxidase
(enzima) atua sobre o peróxido de hidrogênio (substrato) decompondo-o em água e
liberando oxigênio. A orto-dianisidina é uma substância incolor que foi
adicionada à reação para possibilitar a quantificação da atividade enzimática.
À medida que a reação se processa, o corante é oxidado pela liberação de
Oxigênio molecular e vai adquirindo a cor marrom. Desta forma, torna-se
possível determinar por método colorimétrico, empregando-se espectrofotômetro
com polígrafo acoplado, a quantidade de enzima presente na amostra estudada
(Figura 14).

Figura 14- Mary Ann carregando a
prateleira com os tubos de ensaio após ter dosado a HRP por técnica bioquímica.
Notar o tom amarronzado do fluido no interior dos tubos de ensaio.
3. Microscopia
de Luz e Eletrônica de Transmissão
A
absorção de HRP foi também monitorada utilizando-se método citoquímico por meio
da microscopia de luz e eletrônica de transmissão conforme descrição que segue
abaixo.
Após
a conclusão do experimento, o segmento intestinal foi perfundido durante
aproximadamente 3 minutos, a um ritmo de 10 ml/min à temperatura ambiente, com
solução de glutaraldeido 2% tamponada com cacodilato 0,1 (pH 7,2 – 7,4). Após
este tempo os ratos eram sacrificados, o segmento perfundido retirado e
submerso em fixador refrigerado, recentemente preparado. Uma pequena porção
deste material, medindo de 2 a 3 cm de tamanho, distando de 10 a 15 cm da porta
de entrada da cânula proximal era obtido e dividido em pequenos blocos de
tecido de forma cilíndrica. Estes blocos eram submersos em frasco contendo a
solução fixadora e aí permaneciam por uma hora. Após este período de tempo os
fragmentos eram lavados durante 12 horas em solução tampão cacodilato 0,1 M com
7% de sacarose e guardados em refrigerador.
Os
fragmentos cilíndricos eram agora abertos, cortados em porções menores sob
microscópio de dissecção em tampão refrigerado, expondo-se assim a face luminal
da mucosa jejunal. Estes novos fragmentos eram congelados na cabeça de um
micrótomo de congelação, enxaguados durante 15 minutos no meio de Graham e
Karnovsky contendo diamino-benzidina em ausência de substrato (peróxido de
Hidrogênio). Eram posteriormente incubados durante 45 minutos em meio completo
à temperatura ambiente.
O
processo de incubação enzimática era finalizado enxaguando-se brevemente com
solução refrigerada de sacarose 7,5%.
Os
fragmentos incubados eram posteriormente fixados em solução refrigerada de
tetraóxido de ósmio contendo tampão cacodilato 0,1 M (pH 7,3) durante 60
minutos, corados em bloco com acetato de uranila, desidratados em uma série
graduada de soluções de etanol a 50, 70, 95 e 100%, respectivamente, e,
finalmente incluídos em Epon. Cortes espessos de 1 micra eram feitos em ultramicrótomo
para estudo em microscópio de fase (Figura 15).
Figura 15- Microfotografia em
microscópio de contraste de fase mostrando 2 vilosidades jejunais digitiformes
aonde podem ser visualizadas as células epiteliais cilíndricas e as células
califormes. Na superfície mais externa das vilosidades, na região das
microvilosidades, nota-se um contorno enegrecido que corresponde à presença do
produto de reação da HRP (macro-molécula proteica).
Cortes
ultrafinos de coloração prateada eram feitos e examinados em microscópio
eletrônico JEOL-JEM 100. Os espécimes eram estudados levemente corados com
citrato de chumbo. Microfotografias eram obtidas em magnificações iniciais de
5000 a 25000 vezes.
As
análises histológicas dos cortes espessos eram feitas “às cegas”
desconhecendo-se o tipo de tratamento fisiológico ao qual o material havia sido
submetido e interpretadas por 2 investigadores (eu e Saul Teichberg). Nenhum
resultado foi considerado válido caso não fosse reprodutível em pelo menos 3
experimentos independentes. Para cada um dos ratos estudados, 18-24 vilosidades
intestinais por animal foram analisadas em microscopia de fase. Áreas
representativas foram escolhidas para análise em microscopia eletrônica.
Resultados
1. Análise Bioquímica da
Absorção da Horse Radish Peroxidase (HRP)
A
absorção de HRP mostrou-se significantemente mais elevada que
os níveis observados para o grupo controle somente quando a solução de perfusão
continha o sal biliar primário desconjugado (Cólico) e o sal biliar
secundário e desconjugado (Deoxicólico). Outrossim, a absorção
de HRP se revelou mais intensa ainda quando a solução de
perfusão continha o sal biliar secundário e desconjugado. Não houve diferenças
significativas nos níveis séricos da HRP entre os ratos
controles perfundidos com solução glico-salina isenta de sais biliares e nos
ratos perfundidos com o sal biliar primário e conjugado Taurocólico
(fisiológico) (Tabela 1).
2. Microscopia Eletrônica de
Transmissão
A
penetração de HRP através do epitélio intestinal foi citoquimicamente
demonstrável após a perfusão intestinal com o sal biliar primário
desconjugado (Cólico) e com o sal biliar secundário
desconjugado (Deoxicólico). Nestas preparações a HRP foi
visualizada nos espaços intercelulares entre 2 enterócitos adjacentes (Figura
16), bem como ao longo de toda a região das microvilosidades, na zona terminal
no interior de vesículas endocitóticas e corpos multivesiculares, ao longo da
membrana basal das células epiteliais e nos capilares da lâmina própria
(Figuras 17-18).

Figura 16- Ultramicrofotografia de 2
enterócitos adjacentes de uma preparação perfundida com DCh. O produto de
reação da HRP (em negro) é encontrado sobre as microvilosidades (v) e no espaço
intercelular (e). A mitocôndria (m) aparece morfologicamente intacta. No
detalhe, Figura 1a, o produto de reação da HRP (negro) é encontrado ao longo de
todo o espaço intercelular (e) inclusive com ruptura da zona da junção firme
(t). Na Figura 1b o produto de reação da HRP (negro) preserva a região da
junção firme.
Figura 17- Ultramicrofotografia de um
enterócito de uma preparação perfundida com DCh. O produto de reação da HRP
(negro) encontra-se sobre as microvilosidades, as quais estão diminuidas em
altura e ao longo do espaço intercelular. Notar no detalhe que as mitocôndrias
(m) e o aparelho de Gogi (g) encontram-se inchados e deformados.
Figura 18- Ultramicrofotografia de
uma preparação perfundida com DCh. Notar no centro da foto uma estrutura
bizarra arredondada com a presença no seu interior do produto de reação da HRP
(negro). Trata-se de um corpo multivesicular que caracteriza a atividade
lisosomal intracitoplasmática com destruição da macromolécula.
Em
todas as preparações nas quais houve penetração da HRP na
lâmina própria o marcador protéico também foi detectado no interior dos
capilares. O endotélio capilar é fenestrado e a HRP foi
visualizada no interior dos capilares venosos (Figura 19).
Figura 19- Ultramicrofotografia de
uma preparação perfundida com Ch. Nota-se o produto de reação da HRP (negro) no
interior de um capilar venoso da lâmina própria (c), desde o espaça
intercelular (e). Notar em (r) uma hemácia e em (f) um fibroblasto
Com o
sal biliar primário e conjugado, Taurocólico, a absorção de HRP foi
raramente demonstrada, pelo método citoquímico empregado, em apenas umas poucas
vesículas endocitóticas e corpos multivesiculares; o marcador protéico
manteve-se confinado às microvilosidades e ocasionalmente pode-se comprovar
alguns depósitos focais do produto de reação da HRP nas
regiões mais profundas dos espaços intercelulares (Figura 20). Os
mesmos achados também foram observados no grupo controle.
Figura 20-
Ultramicrofotografia de 2 enterócitos adjacentes de uma preparação perfundida
com TCh. O produto de reação da HRP (negro) encontra-se inteiramente confinado
na região das microvilosidades (v). Notar que o espaço intercelular (e)
encontra-se praticamente desprovido do produto de reação da HRP e que as
mitocôndrias (m) e os retículos endoplásmicos (r) encontram-se totalmente
intactos e preservados.





























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}